The evolution and importance of carnivorous plants – uncertain paths of plant carnivory
29 October 2014
When a couple of years ago the first species of carnivorous plants appeared on my window sill – the Venus flytrap and trumpet pitcher – they were only a botanical curiosity and possibly something to scare my guests with. But a single look at them allows the realisation that carnivory itself is not the only trait specific to them. A brief reflection allows for seeing comprehensive adaptations and transformations, of both a morphological and functional nature, manifested in carnivorous plants. For a biologist, the unbelievable precision and concurrence of those adaptations – whose final product is remarkable predacity – provides a matchless example of perfect mechanisms that nature can harness to obtain the best chances of survival in specific conditions.
Currently, in the era of biology based on Darwinian evolution, we look at carnivorous plants from a slightly different perspective. They provide a prime example of how natural selection can direct the development of traits promoting effective growth and reproduction in a specific environment. The biology of carnivorous plants is a perfect illustration of evolutionary compromises behind the life histories of living organisms; exploring their patterns and discovering the forces directing the biological development, we can formulate general conclusions on evolutionary biology and ecology. What’s more – carnivorous plants currently constitute one of the most endangered groups of plants, partly due to their unusual habitat requirements and the resulting vulnerability, and partly due to the progressive degradation of environments inhabited by these species. Thus, an exploration of the biology of carnivorous plants can help in effective protection of these and other endangered species.
The modern diversity of carnivorous plants
Researchers cannot agree on the precise number of known carnivorous plant species.
Currently, almost 600 species of such plants have been described, of which almost all are dicotyledons (only two species belong to monocotyledons). Generally, the carnivorous plants described so far belong to 19 genera grouped into 9 families (Table 1). 13 species appear in Poland: 1 from genus Aldrovanda, 3 from genus Drosera, 3 from genus Pinguicula and 4 from Utricularia.
The uncertainty and variance of these numbers results partly from difficulties in verifying the ecophysiology of particular species.
- must be able to use nutrients obtained from dead animals;
- must have unequivocal adaptations to attract, actively capture and/or actively digest potential prey
Confirming the presence of those traits, particularly the ability to absorb nutrients, is not a trivial task. Carnivorous plants are rather evenly distributed around the world – they appear on all continents (except for Antarctica), both in hot and colder regions. The vast majority of them, however, is limited to quite specific habitats. For reasons described later, carnivorous plants inhabit mostly sunny, humid or even wet (a number of species are water forms) and mostly nutrient-poor (i.e. assimilible nitrogen and phosphorus) habitats. There is also a separate group, characterised with epiphytic growth – consisting in tropical species of Nepenthes pitcher plants and Utricularia bladderworts.
A wide distribution of carnivorous plants is not followed by their local abundance. Generally, the plants are rare, often endangered. Many species are epiphytes. And what is most prominent – despite the sophisticated adaptations of carnivorous plants, there are almost no habitats where they would dominate. The fact seems to contradict the evolutionary level of adaptations that they exhibit, but fits well the theories on evolution of carnivory (presented further below).
Table 1 lists the described genera of carnivorous plants and presents the types of traps used by them. The armoury of traps is admirable and includes most diverse methods of both attracting and killing the prey. What is evident is the great diversity of traps – suggesting that predacity among plants was subject to strong radiation. It should also be noted that carnivory is not a trait characteristic for a single taxon – carnivorous plants are a polyphyletic group of not a systemic but an ecological kind, grouping many lineages.
The phylogenetics and paleobiology of carnivorous plants
The use of molecular methods gave paleobiology and taxonomy new tools in the arduous process of determining evolutionary connections between particular groups of organisms. It is particularly important as it is difficult to establish phylogenetic relations among plants by means of paleobotanics – the plant fossils are rarely found, so the existing fossil record is highly fragmentary.
The interpretation of archaeological data that is commonly accepted nowadays locates the first flower-bearing carnivorous plants in the middle of the Cretaceous period, around 130 million years ago. The oldest preserved fossils include the pollen grains of Aldrovanda genus plants. The oldest and relatively complete fossilised carnivorous plants (Aldrovanda, Drosera) come from the Eocene, which is the period of the Cenozoic starting from which the findings of carnivorous plants become quite regular. An important question arises – did carnivory originate once and the forms that we know today all come from a “common ancestor,” or perhaps it evolved many times over several lineages? If so – are there any “transitional forms” reflecting the acquisition of some carnivore traits, but not its full defining set?
The answer to the first question seems to be settled – the polyphyletic nature of the group formed by carnivorous plants can be interpreted with great likelihood as a result of the development of several parallel evolutionary paths. What is more, their current distribution suggests that in order to reach the current level of biogeographical dispersion, carnivorous plants had to originate in several places in the world. So the existence of a “common ancestor” is not possible.
The origin from a common ancestor would also suggest that specific traits constituting prerequisites of plant predacity were acquired during evolution by a single branch in the phylogenetic (evolutionary) tree and then “radiated” in the process of adaptive radiation. Such an interpretation is highly unlikely, as currently the traits determining this exceptional plant feeding manner are thought not to be evolutionary new (as far as the plant kingdom is concerned). The phenotype connected with plant abilities such as: production of attracting antocyanins and nectars, production of waxes lowering adhesion, presence of proteolytic enzymes and glandular hairs, ability to absorb substances, presence of water “cups” or the motor response ability – is not a new development in evolution; such physiological functions had originated long before as mechanisms underlying other biological processes. Environmental pressure allowed only for their transformation in a specific direction – one can easily imagine a number of parallel processes leading to similar transformation patterns in separate phylogenetic lineages (convergent evolution). It is thus believed that many traits connected with the carnivory of plants are homoplasies — which is also supported by cytomorphological and physiological studies.
The existence of transitional forms is much more likely. Givnish proposed the term “protocarnivory” to refer to species having certain adaptations possibly opening a pathway to carnivory, but not having distinctive traits included in the definition of plant carnivory. Currently it is proposed to apply the term to a couple of species from genus Catopsis.
As Givnish demonstrates, the sequential occurrence of certain adaptations in connection with specific environmental factors (see also further below) can give a plant some abilities whose “side effect” can be carnivory. Consequently, sets of such traits allow for identifying some transitional forms, also in modern flora.
In recent years the paleobotanists’ findings have been confirmed with molecular studies. Comparisons of chloroplast genome sequences (rbcL, MatK, ORF2280) and nucleus sequences (18S rRNA and ITS — Internal Transcribed Spacer) allowed for specifying the relations among carnivorous plants.
Polyphyletism is a fact: the flypaper traps evolved independently at least five times and pitfall traps at least three times. The interesting fact is that phylogenetic genomics allows to demonstrate that even some families are polyphyletic: as it turns out, the Droseraceae family, previously considered monophyletic, contains at least one “odd” taxon (Fig. 1). Consequently, it has been suggested to distinguish a new, separate Drosophyllaceae family. One thing is unanimously proven for certain by molecular data – flypaper traps, so characteristic of Droseraceae, are the most primeval type of catching mechanism and more advanced solutions (e.g. Roridula —> Sarracenia (pitchers); Drosera —> Nepenthes (pitchers); Byblis —> Utricularia (bladders)) can be traced back to them.
Genetic studies also allowed for observation of another intriguing phenomenon – in some phylogenetic lineages originating from initially carnivorous plants, a secondary loss of the ability occurred, e.g. in Ancistrocladaceae and Dioncophyllaceae (excepting the genus Triphyophyllum) – derived from Droseraceae. The network of interconnections in the history of development of carnivorous plants seems complex. What evolutionary traits spurred the rise and betterment of such complex traits?
Carnivory as an adaptation – a winding road to perfection?
Let us imagine a plant in the process of colonizing new habitats. Open spaces and strong sunlight force the secretion of waxes to ensure protection from transpiration. The selection favours individuals having photoprotection elements in the form of higher production of antocyanins. The hollow formed by leaves is filled with rainwater, and odorant and sweet substances accidently fall into it from less resilient leaves. A side effect is the formation of a complex of olfactory and visual signals luring insects to fall into the cup and slide down from the layer of wax. If the plant also has hairs absorbing nutrients, common in many taxa– we get a perfect example of an evolutionary starting pathway to carnivory. Natural selection may favour such individuals, particularly in empty ecological niches corresponding to nutrient-depleted habitats. At this point, carnivory transforms from a trait ensuring an additional source of nitrogen into an adaptation crucial in winning against the ecological competition.
There is, however, an important question – why plant carnivory, which is such an effective adaptation, is also so rare and limited to such unusual species from boggy areas?
An important clue is one of the oldest principles in ecology – Liebig’s law of the minimum. It says that a limiting effect is played by the scarcest environmental factor. We know, however, that the intensity of certain environmental factors has an effect extending to a much more complex level. According to the “life history” approach, which is currently adopted in many ecological studies, the selection of a proper developmental strategy – directed by natural selection – results from evolutionary compromises dictated by various levels of benefits and costs connected with particular choices. In other words, an organism invests its energy in a way ensuring the best possible use of the most limited environmental resources. A plant growing in shade will not invest most nutrients into synthesising new RuBP carboxylase particles, but instead will invest them in photon collecting machinery, as it is the photon flux that is the most limiting habitat factor. Similarly, despite a vast amount of light, desert plants do not develop radiation-absorbing surfaces, instead investing assimilates into development of roots, as they are best suited for habitat limitation posed by a lack of water.
Based on the assumptions connected with evolutionary compromises, Givnish proposed the first model explaining the evolution of carnivory in plants. He considers three potential advantages of carnivory:
- increased pace of photosynthesis is connected with increased intake of nitrogen and phosphorus;
- 2.reproductive advantage connected with storing more nutrient elements in seeds;
- benefit connected with direct replacement of autotrophy with another, organic source of carbon.
Givnish rejects the third possibility outright, postulating that the carbon from killed animals is not assimilated by carnivorous plants. On the other hand, reproductive advantages may result indirectly from the allocation of additional resources to photosynthesis, allowing for a greater reproductive efficiency. It seems that the photosynthetic benefit is the main source of advantages connected with carnivory.
The main assumption in the discussed model is a significant increase in photosynthetic efficiency connected with additional nitrogen and phosphorus supplied thanks to the breakdown of caught prey. Photosynthesis, however, cannot increase indefinitely. In particular – as a photochemical process – its quantum efficiency is directly connected with the flux of light energy received by the leaves in a time unit. At some point, the increase in photosynthesis efficiency should slow down, as other limiting factors come into play. What is more, when the light is poor, the speed of photosynthesis should accelerate significantly more slowly, as the quantity of additional nutrients increase, and that acceleration should plateau more quickly. Consequently, we receive two different photosynthesis curves (Fig. 2) depending on the light and humidity conditions (similarly to light, water also plays a role of a limiting factor in photosynthesis, in connection with transpiration and respiration of photosynthetic organs). A simple comparison of the benefits and costs shows that in good light conditions and abundance of water, the availability of nutrients must be the limiting factor. In other words: when light and water is abundant, the difference between profits and costs (net profit — dashed lines in Fig. 2) exceeds the cost of retaining carnivory near C=0 (the fragment where the slope of curve P — C is greater than zero).
Indirect evidence for the operation of such an evolutionary mechanism is so-called heterophylly in carnivorous plants, manifesting itself with the production of leaves without traps or with vestigial traps if the conditions are unfavourable (e.g. drought, low temperature, seasonal underexposure to light). This phenomenon is probably directly connected with an increase in the costs of carnivory – e.g. when water is missing, the cost of producing digestive secretions may be too high and the plant simply gives up producing catching leaves to relieve the stress. Heterophylly is known to occur in many genera, including Dionaea, Sarracenia, Cephalotus and Nepenthes. The genera not showing such adaptations usually use other forms of phenotypic plasticity. Species from the Pinguicula and Drosera genera increase gland density and production of mucilaginous (sticky) secretion only upon stimulation, i.e. catching the prey – thus reducing the cost of “idle” secretion of killing substances.
The discussed model is also referred to in attempts to explain the rarity of carnivorous plants. The right combination of lighting and hydrological conditions is not frequent (e.g. wet habitats – like bogs and tropical forests – are also most frequently very shady). Last but not least, also the most important limiting factor must be present, i.e. infertile soil. If such niches are available – they will almost certainly be inhabited by carnivorous plants. Most frequently, however, such niches of possible carnivorous plant species’ development are only small patches in vast areas – anywhere else their adaptation is too costly and N autotrophic species are favoured.
Although the Givnish model seems to be a good approach to explaining the evolution of carnivory in plants – it is not without its defects. The most important objection may probably concern its excessive simplification. Ecological regularities depend on the complex relations of many parameters. Therefore, it seems that not only moisture and light should be taken into account, but other factors as well (especially since those two parameters are not the only ones decelerating photosynthesis). Objections can also be raised against one of the model’s assumptions that the plants do not assimilate carbon obtained from prey (which would decrease photosynthetic efficiency in connection with partial carbon heterotrophy). It does not seem particularly likely for carnivorous plants not to evolutionarily develop the ability to utilise carbon (which could constitute a very important evolutionary advantage), especially since the absorbed compounds include not only ammonium nitrogen, but also amino acids – composed, after all, of nitrogen, carbon and oxygen.
Their presence has been discovered in the liquid of many pitcher traps, and absorption was confirmed by isotropic studies using 3H-labeled derivatives. Similar isotopic studies should therefore be conducted taking into account the partitioning of carbon coming from animals in plant cells. The most serious objections against the Givnish model, however, come from experimental studies.
Although the photosynthetic activity that changes the formula proposed by him conforms to many observations, there is also evidence available for completely contradictory correlations. Specifically, in several species (mostly subarctic) a negative correlation was found between photosynthesis efficiency and the intensity of predacity. Attempts have been made to explain the observations by reference to other plant resource allocation patterns. Perhaps the nitrogen is used for synthesising metabolites not related to photosynthesis? Or perhaps – which some results seem to suggest – Givnish’s assumption on the lack of direct contribution of the absorbed nitrogen to reproduction (apart from photosynthetic efficiency) also requires revision? Finally – perhaps carnivory changes the anatomy of photosynthetic organs (especially the portion of efficient palisade cells) so significantly that it affects photosynthetic efficiency.
In the light of the quoted arguments, it seems obvious that Givnish’s model – however interesting thanks to the “profit-cost” approach – is only a starting point for move complex solutions based on the ecological niche concept.
One such model is Benzig’s proposal based on a three-dimensional niche defined by three parameters: sunlight, humidity and abundance of non-organic matter. Benzig’s inspiration were plants from the Bromeliaceae family, where carnivory is relatively rare (2 species), but other forms of adaptation to poor habitats are present: use of dead organic matter falling into cups formed by leaves and myrmecophily (association with ants providing nutrients and “tending” in exchange for shelter in the plant structure). What differentiates Benzig’s model from Givnish’s proposal is the possibility of carnivory occurrence in a number of light/water regimes, which results from the presence of compromises between a given factor and the other two parameters. In other words – the sets of parameters create spaces in the niche where occurrence of a given type of feeding is possible from the evolutionary perspective, and a change in those parameters may constitute a kind of a “switch” moving the strategy from one area to another (Fig. 3). This system of selection factors allows the Benzig’s model to best explain the occurrence of epiphytic forms of carnivorous plants in the environment of shady tropical forests.
The referred models allow for foreseeing whether carnivorous plants in certain conditions can gain an adaptive advantage and, thus, whether this feature can be maintained as a stable feature. Yet, what does this adaptive advantage consist of?
How does it manifest itself within a specimen’s physiology? A number of experiments performed on many varieties of carnivorous plants prove their adaptation – measured as reproductive potential, vegetative reproduction or vegetative growth intensity – to increase if artificial feeding with additional prey is applied. The plants responded with an increase of size, blossom intensity and investment in predatory mechanisms. Therefore, carnivorous plants rely on biogens obtained from digesting their prey, whose abundance in the environment is an important limiting factor of the plants’ growth and reproduction.
Interestingly, the extent of animal nitrogen usage differs among 28 groups of predatory plants. According to research on 15N nitrogen distribution in tissues, the usage of animal nitrogen differs even within a given specimen. Low 615N values (the amount of nitrogen-15 isotope related to the reference group), characteristic for soil nitrogen, are typical in older prehensile organs (as proven in Nepenthes and Cephalotus). The majority of animal nitrogen escapes from those organs (nitrogen sources) and accumulates in younger organs (nitrogen sinks), thus increasing the amount of 615N in them. In genus Cephalotus, the overall amount of N increased also along with the number of open pitchers. Moreover, for this genus it was blossoming that depended most on the amount of animal nitrogen: flowers contained the most of the animal nitrogen. This differentiation occurs at the intergenetic level as well: the highest content of animal nitrogen is evidenced in species with the most complex prehensile systems (e.g. Drosera – average of 30-40%, whereas Nepenthes or Heliamphora – about 70-80%). These observations prove the dependence patterns of plants on additional nitrogen sources to be different, and the content gradients to dynamically change within single specimens, being strongly dependent on the history of individual development. The overall picture is even more complicated due to differentiated nitrogen allocation in structures directly benefiting from plant carnivory. A number of species are investing surplus resources in vegetative development and an improvement of prey catching effectiveness. Yet there are species producing spore structures, like winter buds, during their individual development and redirect there the majority of resources derived from carnivore activity.
This phenomenon, referred to as preformation, is a real challenge in ecological research, as the “escape” of biogens to spore structures can cause even a several months delay in a plant’s response to experimental procedure (like additional feeding), which makes the assessment of results difficult.
Adaptation to carnivory – not always as perfect…
It seems rather clear that being a carnivore requires specific adaptive measures. The array of ways of killing, unique and reserved only for this ecological group of plants, is indeed impressive. Glue, bladder, and hamper traps – this is only the beginning of the list. Just as refined are pools containing digestive liquid able to decompose even small vertebrates (by the way, the remains of small vertebrates and hummingbirds found in Nepenthes’ pitchers were the reason for changing the name “insectivorous plants” to “carnivorous plants”). The deadly engineering culminates in snap traps of the Dionaea muscipula species – including a number of mechanisms not only instantly reacting to the presence of prey, but also preventing from “false alarms” or false signals about a prey’s presence, while in fact it was not caught.
It turns out, however, that adaptation to carnivory is not always as effective. One of the earliest recognised evolutionary conflicts is the interaction of the traps with the reproductive processes of the plants. It is mainly expressed in the casual killing of pollinators, which results in a striking conflict of interest in plants attempting to gain additional resources, while on the other hand to invest them in effectiveness of reproduction.
The species researched the most from this perspective is the endemic butterwort, Pinguicula vallisnerifolia. The experiments consisting in excluding certain species of insects from this plant’s environment confirm that its reproduction is highly limited by the presence of pollinators, particularly large species capable of cross-pollination, which is the most effective in terms of quantity and durability of produced seed. On the other hand, a significant correlation was proven between the number of insects found in the flowers and being caught by the leaves. Both groups of insects contained identical species, with the particular presence of pollinating species among the prey. Thus, the pollinator-prey conflict is quite visible. Moreover, its intensity is modified by environmental factor gradients. In very sunny areas, where large insects prevail like butterflies and large beetles (the effective cross-pollinators), which are harder to catch, the conflict is not as visible. Mainly small insects are caught (small beetles, thirps, mainly self-pollinating), which in fact paradoxically increases the level of adaptation of the plant by reducing the participation of those less effective pollinators (thus reducing the probability of self-pollination). Additionally, the plant regains some of its investment in pollen by partially digesting it along with the insects killed. In shady habitats there are much fewer large insects: here the main pollinating force are smaller insects and their being accidentally caught reduces the plant’s adaptation – thus aggravating the conflict.
This conflict manifests itself even more clearly when it comes to plants with pitcher traps.
Due to convergent evolution, these pitchers are equipped with attracting mechanisms similar to those found in flowers.
These are mainly the smell and the taste of nectar. Some pitcher plants and trumpet pitchers bear colourful patterns on their traps, being precise signposts for insects. As it turned out, the spectral characteristics of those patterns (especially in UV light) perfectly correspond to the wavelengths to which insects’ sight is most sensitive – there is no doubt regarding the purpose of such features. Experiments consisting in masking the patterns or artificially manipulating the smell give results compliant with the expectations i.e. a rapid decrease of effectiveness in catching animals right after a given feature was removed.
The decrease in adaptation should, however, result in a specific evolutionary effect. Many species of carnivorous plants have developed mechanisms that reduce the intensity of the pollinator-prey conflict. It happens mostly due to temporal or spatial separation of prehensile organs and flowers (Fig. 4), which prevents the capture of pollinating species. In some trumpet pitchers and pitcher plants, the strategy is quite different: they have evolved to send specialised visual and olfactory signals to an extent that they only attract prey, at the same time not being attractive to the pollinators.
Manipulation of visual and olfactory signals can also have a completely different function. Just like catching potential pollinators, also excessive catching of any prey may be harmful: overloaded traps may result in rotting and thus put the entire plant at risk of death. The specialisation – and sometimes even the weakening of certain signals – may therefore have a great significance for regulating the amount of caught prey. Some researchers also underline certain behavioural aspects. “Passing” some prey may be important for spreading information on the potential source of nourishment (which is a well-known phenomenon in the case of social Hymenoptera). Such a behaviour of insects would then have a favourable influence on the later “hunting” success for a given specimen. It turns out that, in some cases (genus Nepenthes), the ground pitchers, which have a much greater chance of catching the prey, have their attracting elements significantly reduced.
Carnivorous plants in the context of their ecosystem
No matter how rare and unique an organism we consider, it never lives all by itself. The functioning of every species can be considered on many levels; but only the ecosystem level, which engages interactions with other organisms, is the most complete one, also from an evolutionary point of view. Carnivorous plants, thanks to their exceptional adaptations, can interact with the environment in many ways.
The interaction potential is immense. The fact itself that these species take a direct part in the organic matter cycle both as producers and consumers is already intriguing. Interestingly, there are no research programmes aimed at evaluating the participation and significance of this group of plants in the flow of energy in the ecosystem. This is partially due to the fact that these plants are rather rare: they hardly ever dominate plant communities. Furthermore, such research would be extremely difficult and, which may be even more important, costly to perform, due to the necessity of engaging expensive isotopic preparations.
Nevertheless, we seem to already have an insight into these plants’ place in ecosystems. There is no doubt that their importance reaches far beyond mere processing of their prey. Digestive fluids of many insectivorous plants with pitcher traps contain bacterial cultures that bind nitrogen. Therefore, they are an essential source of nitrogen – especially in places where the soil is poor in this biogen (tropical laterite soils).
Also, complex dependencies between predatory plants and animals should not escape our notice; and it is not just about a mere antagonistic interaction of predators. One of the most surprising findings was to discover carnivorous species that… converted to vegetarianism! Researchers found plants which, instead of catching insects, would allow those insects to be eaten by tropical poison dart frogs and small birds. In exchange for attracting their prey, the vertebrates defecate onto their partner plants’ pitchers, thus providing a constant and far easier absorbable source of nitrogen.
Some species of genus Nepenthes were also discovered which, growing on the ground, allowed their pitchers to grow into the surface. As a result, some plant remains from the surrounding groundcover fall into the pitcher and are decomposed.
Vegetarianism – what else?
There are also some species worth mentioning as, although they did not give up on their animal prey, they changed their hunting method. Many species producing nectar entered into mutualistic interactions with small insects.
The latter feed on the plant’s secretions, at the same time playing the role of so-called secondary attractants, luring much bigger and more valuable prey right into the starving traps.
Add to it the distribution of minor fauna species in the habitat, which is typically related to the specialisation of particular species in catching specific prey – and we have a group of organisms playing a vital ecological role. We still seem to underestimate their significance; yet we should bear one thing in mind. The habitats hosting carnivorous plants are dwindling at an alarming rate. Unless serious measures are taken, this beautiful and exceptional group of plants may disappear from our planet’s surface. This would mean having lost not only the species themselves but also the amazing history of evolution standing behind their origin.
Author: Szymon Drobniak
Sources:
1. Benzing D.H. 1987 — The origin and rarity of botanical carnivory — Trends Eco.Evo. 2: 364-369
2. Ellison A.M., Gotelli N.J. 2001 — Evolutionary ecology of carnivorous plants — Trends Eco. Evo. 16: 623-628
3. Givinish J.T., Burkhardt E.L., Happel R.E., Weintraub J.D. 1984 — Carnivory in the Bromeliad Brocchinia reducta, with a cost/benefit model for the general restriction of carnivorous plants to sunny, moist, nutrient-poor habitats — Am.Nat. 124: 479-497
4. Mćndez M., Karlsson P.S. 1999 — Costs and benefits of carnivory in plants: insights from the photosynthetic performance of four carnivorous plants in a subarctic environment — Oikos 86: 105-112
5. Moran J.A. 1996 — Prey attraction in N. rafflesiana — J. Ecol. 84: 515-525
6. Moran J.A., Booth W.E., Charles J.K. 1999 — Aspects of pitcher morphology and spectral characteristics of six Bornean Nephentes pitcher plant species: implications for prey capture — Ann. Bot. 83: 521-528
7. Pietropaolo J., Pietropaolo P. 1997 — Carnivorous plants of the world — Timber Press, Inc. (Portland, USA)
8. Schulze W., Schulze E.D., Pate J.S., Gillison A.N. 1997 — The nitrogen supply from soils and insects of the pitcher plants Nephentes mirabilis, Cephalotus follicularis, and Darlingtonia californica — Oecologia 112: 464-471
9. Thorćn L.N., Karlsson P.S. 1998 — Effects of supplementary feeding on growth and reproduction of three carnivorous plant species in a subarctic environment — J.Ecol. 86: 501-510
10. WorleyA.C., Harder L.D. 1999 — Consequences of preformation for dynamic resource allocation by a carnivorous herb Pinguicula vulgaris (Lentibulariaceae)
11. Zamora R., Gómez J.M., Hódar J.A. 1997 — Responses of a carnivorous plant to prey and inorganic nutrients in a Mediterranean environment — Oecologia 111: 443-451
12. Zamora R., Gómez J.M., Hódar J.A. 1998 — Fitness responses of a carnivorous plant In contrasting ecological scen arios — Ecology 79: 1630- 1644
13. Zamora R. 1999 — Conditional outcomes of interactions: the pollinator-prey conflict of an ainsectivorous plant — Ecology 80: 786-795
Table 1: Overview of taxa for carnivorous plants (the corresponding names in English are given in brackets after the Latin names)
| Order | Family | Genus | W* | Ł* | T* | Trap type | Num. of species† |
|---|---|---|---|---|---|---|---|
| Sarraceniales | Sarraceniaceae | Darlingtonia | + | – | – | Pitcher | 1 |
| Heliamphora | + | – | – | Pitcher with bait | 3+ | ||
| Sarracenia (kapturnica) | + | – | ± | Pitcher | 9x | ||
| Nephentales | Nephentaceae | Nephentes | + | – | + | Pitcher | 80+x |
| Droseraceae | Aldrowanda | – | + | + | Snap trap | 1 | |
| Drosera (Sunde) | – | + | + | Active sticky tentacles | 90+x | ||
| Dionaea (Venus flytrap) | + | + | + | Snap trap | 1 | ||
| Drosophyllum (Dewy pine) | + | + | + | Sticky tentacles | 1 | ||
| Violales (Fiołkowce) | Dioncophyllaceae | Triphyophyllum | – | + | + | Sticky tentacles | 1 |
| Scrophulariales | Lentibulariaceae | Utricularia (Bladderwort) | – | + | + | Prehensile bubbles | 250+ |
| Polypompholyx | – | + | + | Prehensile bubbles | 2+ | ||
| Gelinsea (żelinsea) | – | – | + | Funnel basket trap | 12 | ||
| Biovularia | – | – | + | Bubbles | 2 | ||
| Pinguicula (tłustosz) | – | – | + | Sticky leaf surface | 30+x | ||
| Saxifragales (Skalnic owce) | Byblidaceae | Byblis | – | – | + | Sticky tentacles | 2 |
| Roridulaceae | Roridula | ? | – | + | Sticky tentacles | 1+ | |
| Cephalotaceae | Cephalotus | + | – | + | Pitcher | 1 | |
| Bromeliales | Bromeliaceae | Brocchinia | + | – | + | Dish-shaped leaves | 1 |
| Catopsis | ? | – | – | Dish-shaped leaves | 1 |
A – the presence of prey-attracting mechanisms; C – active catching mechanisms (‘-’ stands for a passive mechanism); D – the presence of proteolytic enzymes for digesting the prey (‘-’ – digestion with microorganisms)
† The ‘+’ sign means a number of given species was specified, although some sources feature higher figures; the ‘x’ sign means that there are many hybrid forms known (mainly breading morphs).
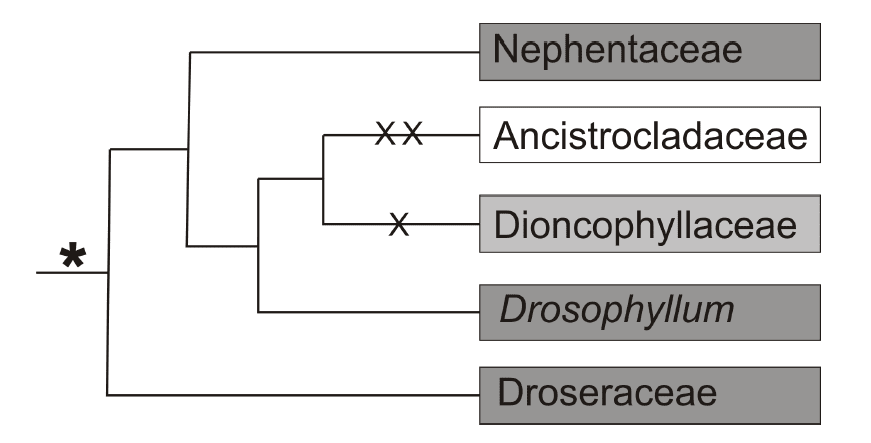
Fig. 1: The phylogenetics of a couple of families of insectivorous plants; an asterisk – carnivory as an original feature; X – loss of carnivory in several genera; XX – total loss of carnivory.
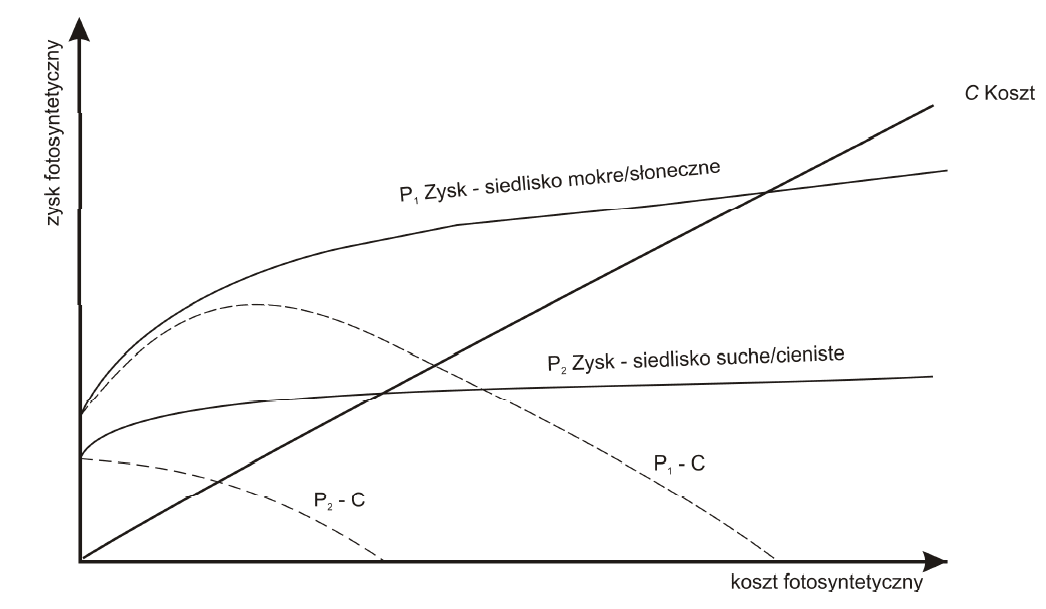
Fig. 2: The Givnish model for the evolution of carnivory in plants. (according to Givnish et al. (1987))
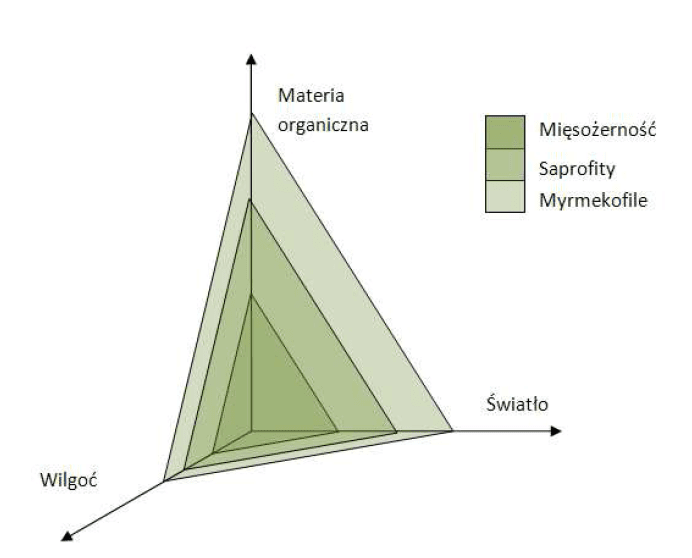
Fig. 3: The Benzig model for the evolution of carnivory; the surfaces limit the areas of occurrence of suitable feeding strategies.
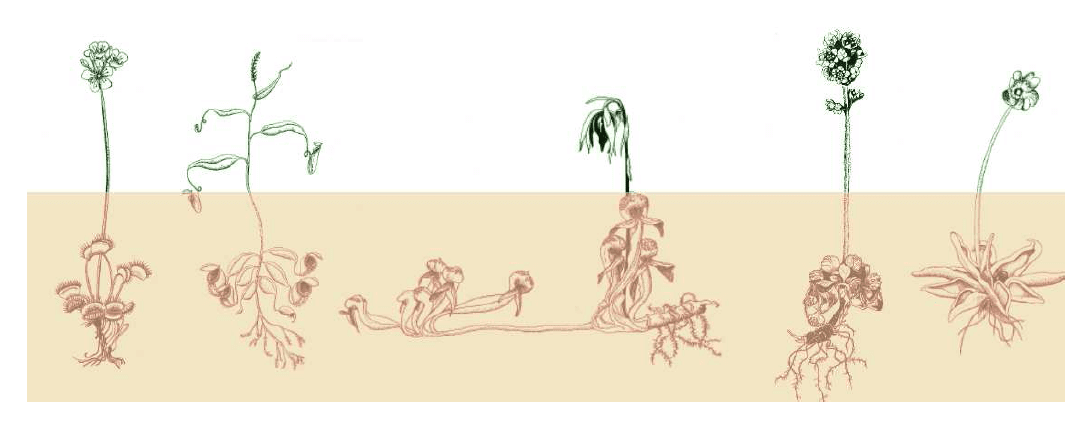
Fig. 4: Spatial separation of prehensile organs (red) and flowers as a safeguard against the pollinator-prey conflict; from the left: Dionaea, Nepenthes, Darlingtonia, Cephalotus, Pinguicula.















